
Controle Motor
O controle dos movimentos do nosso organismo podem ser divididos em três categorias: pode ser voluntário, involuntário ou controle automático (classificação intermediária). As diferenças estão fundamentalmente na região hierárquica superior de controle.
Controle Voluntário
O movimento voluntário baseia-se na produção e execução de um comando de forma consciente e controlável (modulável). Para tanto, o movimento precisa ser organizado inicialmente ao nível de córtex sensório-motor. Deste modo, o evento da contração muscular antecede o movimento, ou seja, antes do movimento propriamente dito começar, já há ativação muscular (encurtamento dos sarcômeros). De fato, há necessidade de geração de tensão suficiente para movimentar carga ou vencer resistência. O córtex motor, mais especificamente, organiza o comando do movimento e envia este comando para um nível/centro inferior, numa seqüência relativamente bem definida: córtex sensório-motor, tálamo, gânglios da base, tronco encefálico, medula, músculo (proprioceptores).
Feedback Muscular: A informação a respeito do resultado da contração inicial ascende ao cerebelo, onde há a comparação entre o movimento realizado e o idealizado, que nunca são iguais. Esta diferença se deve às variáveis ambientais, como a força peso, resistência da vestimenta, forças de atrito ou arrasto, entre outras, variáveis que afetam o desempenho do gesto motor. O resultado é a discrepância, em níveis variados, entre o movimento que foi programado pelo córtex sensório-motor e o movimento que foi efetuado pelo tecido muscular. O cerebelo, então, informa o resultado desta comparação, enviando as devidas correções de volta para o córtex sensório-motor, de onde partem inúmeras microcorreções à medida que o movimento transcorre. No caso de uma lesão cerebelar, por exemplo, as correções devem ser feitas de forma consciente, ficando os erros muitíssimo mais evidentes. Tais erros incluem força excessiva, força insuficiente, mudanças indesejadas de direção, entre outros problemas que não se pode prever. Sem atividade cerebelar adequada, não há controle fino do movimento (este controle, na verdade, é executado de forma involuntária ou inconsciente). Quanto mais estranho, incomum, diferente ou novo for o movimento, maior a participação do cerebelo corrigindo a trajetória, desde o início da contração até o alcance do alvo/objetivo.

Os engramas são alguns gestos que já estão aprendidos e são amplamente treinados, executados e repetidos como caminhar, escrever, falar, etc. A fim de efetuar tais ações, não há mais a necessidade de pensamento preciso e estudado, pois eles já são realizados de forma automática. Para movimentos novos e para gestos que não são habitualmente repetidos, a organização motora é totalmente voluntária, necessariamente.
Durante os primeiros anos do processo de desenvolvimento motor e neural, a seqüência lógica e precisa de ativação neuronal visando a execução de movimentos voluntários não necessariamente ocorre de forma correta ou seguindo o caminho mais curto. O produto final desta transmissão confusa de sinal é a ativação inadequada da musculatura solicitada para o movimento, a contração simultânea de uma série de músculos não envolvidos no objetivo, entre outras deficiências. O processo de repetição que possibilita a aprendizagem é uma forma de exercício da via neuronal, perdurando até que o caminho correto, desde o córtex até o músculo, seja reconhecido e posteriormente gravado. Ao longo do tempo, à medida que o treinamento motor transcorre, forma-se um padrão de movimento. Num dado momento do desenvolvimento, cuja determinação precisa é bastante difícil, o exercício funcional continuado faz com que a via seja simplificada, e o centro de comando de controle motor migra do córtex sensório-motor para o tronco cerebral. O córtex sensório-motor continua sendo fundamental para dar início ao movimento que, afinal, é voluntário; assim que ele é iniciado, no entanto, a regência do movimento passa para a responsabilidade do tronco cerebral, de forma que marcha-se, dirige-se, mastiga-se, escreve-se, fala-se, entre outras ações, sem a necessidade consciente de manutenção do gesto. Em outras palavras, o movimento passa a ser automatizado, que é um estágio intermediário entre os movimentos voluntários e involuntários.
Da mesma forma que o gesto motor é estruturado e automatizado, as características individuais, bem como erros eventuais, também o são, e o padrão motor memorizado é definido ao nível neuromuscular a ponto de ser extremamente difícil a sua alteração ou correção. Muito embora seja bastante difícil modificar padrões motores estabelecidos, ainda há a possibilidade de correções em graus variados. Por isto, atividades como a marcha, a escrita e a fala, entre outras, são de fato peculiares a cada indivíduo. Situações patológicas ¾ como o acidente vascular cerebral ¾ podem fazer com que o indivíduo perca o padrão motor adquirido, em função da lesão de determinada região do córtex motor. Nesta situação, os tecidos periféricos à área atingida (área circunjacente) podem assumir o controle dos movimentos anteriormente organizados pela área agora necrosada, possibilitando ao indivíduo afetado o reaprendizado dos gestos inicialmente estruturados.
Engramas Acessórios
Existem alguns gestos que não estão necessariamente relacionados à dinâmica seqüencial básica de execução motora, ou seja, ao engrama original. Tais movimentos são decorrentes do refinamento no aprendizado e relacionam-se à estruturação neuromuscular de padrões motores. Nesta dinâmica da formação de engrama, indivíduos com maior experiência motora (vocabulário ou repertório motor), apresentam uma capacidade de improvisação infinitamente superior a indivíduos restritos às ações cotidianas. Este fato decorre da capacidade de transferência da vivência motora entre várias situações. Um atleta do handebol, por exemplo, pode ter, em princípio, muito mais facilidade em adaptar-se aos gestos do vôlei. Esses engramas são normalmente associados à bagagem de movimentos que o indivíduo traz desde o início da sua formação.
Controle Involuntário
Existem basicamente 3 reflexos medulares, que são reflexos organizados e efetuados sem a participação de centros superiores:
- Reflexo miotático;
- Reflexo miotático inverso;
- Reflexo de retirada.
O reflexo miotático baseia-se nas informações fornecidas pelo órgão sensorial denominado fuso muscular. O fuso é formado por células musculares bastante diferenciadas que não possuem sarcômeros na região central (apenas as extremidades são contráteis); junto ao equador celular, encontram-se os múltiplos núcleos (centenas) da fibra muscular esquelética; ao redor da região central, encontra-se uma fibra nervosa aferente do tipo IA espiralada, que responde à deformação mecânica; as extremidades do fuso muscular são inervadas pelo motoneurônio gama, enquanto as fibras musculares extrafusais ou regulares são inervadas pelo motoneurônio alfa. O reflexo miotático é o único reflexo monossináptico humano, ou seja, a conexão entre as raízes sensitiva e motora se faz diretamente no interior do H medular, sem mediação por interneurônios.
A fibra fusomotora serve para modular o comprimento da fibra intrafusal, mantendo-a como um sensor ativo em qualquer estado de comprimento muscular. Quando há encurtamento muscular ¾ mesmo na ausência de contração ativa, ou seja, devido ao encurtamento passivo por aproximação de origem e inserção musculares ¾, há ativação motora gama por via reflexa, a fim de evitar a deformação das fibras intrafusais, que estão em paralelo às fibras extrafusais, mantendo sua função sensorial. A contração das extremidades do fuso levam à distensão do centro fusal, ao redor do qual está a terminação nervosa sensitiva. Caso o sistema gama não apresentasse atividade, o encurtamento muscular levaria à perda da sensibilidade de comprimento do músculo.
O reflexo miotático, então, decorre da estimulação do aferente IA devido ao estiramento muscular, seguida do envio do sinal adequado para a medula, onde, através de uma única sinapse, o neurônio motor ado agonista é estimulado, gerando contração muscular reflexa, e o neurônio motor a do antagonista é inibido, mantendo-o relaxado.
O reflexo miotático é exigido em situações de recepção (amortecimento) de projéteis, por exemplo, quando a força de impacto causada pelo contato do objeto gera o estiramento muscular, que ativa a circuitária nervosa, causando a contração do tecido muscular estirado. As perturbações mecânicas sobre tendões geram estiramentos musculares de pequena amplitude, que são percebidos pelas fibras intrafusais, gerando respostas reflexas. Apesar da localização do OTG ser tendínea, o seu limiar de excitabilidade é muito mais alto, e sua ativação deve ser realizada por mais tempo (fala-se até em 6 segundos) a fim de que um potencial de ação seja desencadeado.
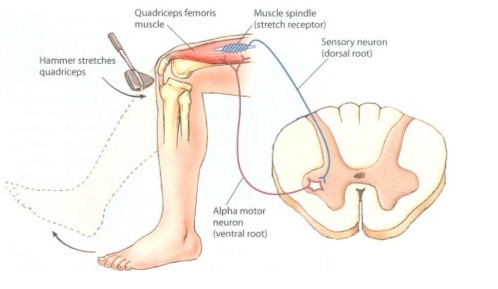
Este reflexo também é fundamental para a manutenção da postura e resistência à ação da força gravitacional. Durante sessões de treinamento pliométrico, atletas de inúmeras modalidades esportivas executam, por exemplo, saltos a partir de superfícies mais altas, com alto impacto, seguidos de saltos sucessivos para áreas adjacentes, elevadas ou não. O primeiro contato do segmento com o solo se dá através da ponta dos pés, seguido pela flexão dorsal, flexão de joelho, flexão da articulação coxo-femoral; assim que o movimento tenha sido transmitido ao longo de todas as articulações do segmento, a contração voluntária é começada. O objetivo do treinamento pliométrico é potencializar a contração muscular voluntária, somando-a à resposta reflexa desencadeada pelo estiramento da musculatura. A sobreposição das ações voluntária e reflexa, requerida através da ativação miotática no momento do impacto, possibilita maior recrutamento de unidades motoras e um incremento significativo no processo de coordenação neuromuscular. A pliometria se aplica também a membros superiores, além de permitir variações na execução e adaptação da técnica (saltos com um pé só, alturas variáveis, entre outras possibilidades). Treinamentos de membros superiores podem ser realizados, por exemplo, através da manipulação (interceptação ou recepção) de medicineball (indivíduo apoiado contra parede ou deitado no chão); o processo de amortecimento é o responsável pela ativação do reflexo miotático; segue-se a este a ação voluntária de lançar a bola de acordo com o gesto efetuado no esporte praticado. Ainda que o atleta não reproduza esta situação nas competições, o recrutamento é aumentado em função das duas vias utilizadas: a voluntária e a involuntária, e o aprendizado da coordenação neuromuscular para o recrutamento se mantém. A longo prazo, em termos de valências físicas, ganha-se fundamentalmente potência para o gesto a ser realizado. O reflexo miotático também é exigido em inúmeras outras situações cotidianas ¾ quando o indivíduo dorme sentado, por exemplo. A indução do sono faz com que a musculatura relaxe e a cabeça penda, estirando o tecido muscular e a fibra intrafusal; este alongamento deforma o aferente IA, ativa a contração reflexa e, provavelmente, leva o indivíduo a despertar (o padrão encefalográfico entre o sono e a vigília comporta-se como um processo de liga e desliga, ou seja, não há aviso prévio ou alteração gradual; simplesmente passa-se do sono para o estado desperto e vice-versa).
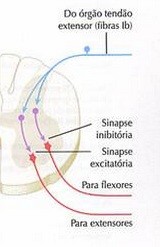
O reflexo miotático inverso, baseia-se na ativação do Órgão Tendinoso de Golgi, que é um sensor responsivo à contração muscular. O OTG é uma estrutura (terminação nervosa livre) encapsulada, localizada na junção músculo-tendinosa e que está em série em relação ao tecido muscular.
Em conseqüência de sua disposição anatômica, o OTG não é capaz de informar a respeito da variação de comprimento muscular (se o músculo encurta, ele sofre tração; se o músculo alonga, ele também sofre tração). Os OTGs são designados como sensores de força, ou seja, estruturas responsáveis por informar ao SNC a magnitude do estado contrátil do músculo em cujo tendão está inserido. O estímulo que causa a ativação do Órgão Tendinoso de Golgi é a tensão transmitida ao osso pelo encurtamento dos sarcômeros; ele é insensível a variações no comprimento muscular. Diante da contração muscular, estes sensores enviam sinais para o SNC através da fibra aferente IB. Esta fibra penetra na região posterior da medula e conecta-se a dois interneurônios: um excitatório, que transmite o impulso ao eferente do antagonista, e um inibitório, que bloqueia o sinal para a fibra motora do agonista, causando seu relaxamento máximo e impedindo sua contração por décimos de segundo. Assim que a tensão desaparece, o reflexo é interrompido, habilitando o músculo para novas ativações (contrações).
O estímulo (tensão) para o desencadeamento da atividade do OTG, segundo dados da literatura, deve perdurar por 6 segundos. A contração do antagonista simultaneamente ao relaxamento do agonista tem por função evitar o processo de estiramento. O reflexo miotático inverso está relacionado à proteção das estruturas relacionadas ao sistema músculo-esqueletal e à produção de força e movimento. Assim, se a tensão gerada atingir um nível crítico, potencialmente lesivo aos tecidos relacionados, incluindo a possibilidade de rompimento tecidual, o reflexo miotático inverso causa o relaxamento da musculatura que sustenta a carga, interrompendo o processo de contração muscular e produção de força. A queda-de-braço é um bom exemplo da atuação protetorado reflexo miotático inverso, através da ativação do OTG. Se um indivíduo tenta aparar ou segurar um objeto demasiadamente pesado, o primeiro reflexo ativado é o miotático, já que o primeiro estímulo é o estiramento, informando ao SNC o número de unidades motoras necessárias à realização do movimento e a magnitude de contração; a seguir, diante da constatação do dano iminente, o reflexo miotático inverso é ativado. Em outras palavras, a contração proporcional à carga elevada gera um grande aumento na tensão sobre o Órgão Tendinoso de Golgi. Enquanto o reflexo miotático inverso protege os tecidos do rompimento, o reflexo miotático é um mecanismo de defesa contra alterações posturais. Situações extremas, envolvendo fratura por tração, ocorrem mais comumente em casos de osteoporose avançada. Apesar da musculatura estar também debilitada, a resistência à tração do osso está ainda mais diminuída. Fraturas que afetam os atletas das mais variadas modalidades em geral decorrem de forças de impacto, que não tem relação com o OTG.
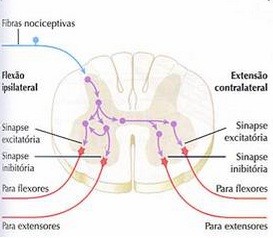
O reflexo flexor de retirada, é mediado por um arco-reflexo polissináptico, no qual estímulos nociceptivos e termoceptivos são impulsionados por neurônios aferentes e mediados por neurônios de associação que estimulam neurônios de um ou mais segmentos medulares para executar a flexão do membro estimulado. Paralelamente, os mesmos neurônios aferentes emitem colaterais que cruzam a linha média da medula e excitam os motoneurônios alfa. Estes se dirigem aos músculos extensores do membro contralateral. Um exemplo é quanto pisamos em um prego, ou colocamos uma mão em uma chapa quente.
ALONGAMENTOS:
O alongamento é um método frequentemente executado para aumentar a amplitude de movimento ou a flexibilidade de determinadas articulações. Ao contrário do que muitas pessoas acreditam, os métodos de alongamento não são eficientes em relação à prevenção de lesões á curto prazo. As técnicas de alongamento são empregadas com três principais finalidades:
- Ganho de flexibilidade -> Envolve:
- Elasticidade da pele;
- Massa adiposa/muscular;
- Elasticidade muscular (depende da viscosidade do sarcoplasma) -> treinamento gera aumento do número de sarcômeros em série;
- Mobilidade articular;
![]()
- Relaxamento muscular
- Aquecimento:
![]()
Existem três tipos de alongamentos que são mais utilizados:
-
Alongamento Balístico: é o tipo de alongamento que está associado a movimentos insistentes, como por exemplo, pendulares, saltos e movimentos rítmicos.
-
Alongamento Estático: é aquele onde o membro deve ser levado até sua amplitude máxima, e ali permanecer durante o período estipulado para a realização do alongamento. Deve ser aplicado de forma lenta e gradual para que não ocorra o reflexo de estiramento e a estimulação do OTG.
-
Alongamento Facilitação Neuromuscular Proprioceptiva (FNP): é o método baseado na potencialização do reflexo miotático inverso (OTG) e na inibição do reflexo miotático (fuso muscular). Esta técnica é aplicada por um profissional, que leva o membro até sua amplitude máxima e pede uma contração, aplicando uma resistência. Após a contração isométrica, o paciente relaxa e o profissional consegue levar o membro à uma amplitude do que a inicial.
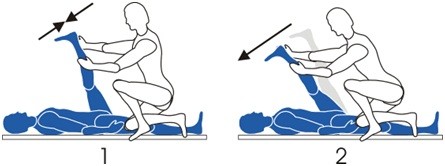
1. Paciente em decúbito dorsal e com quadril flexionado até sua amplitude máxima. O profissional pede a contração e resiste ao movimento.
2. O paciente relaxa o membro e o profissional consegue ir além da amplitude inicial.
Resumindo...
